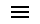

黄种人优势与缺点(黄种人的三大分支情况如何)
黄种人优势与缺点(黄种人的三大分支情况如何)
黄种人O演化为三大支系
3万年前,NO人群分化成N人群和O人群,此后N人群的一部分成功北上,也有可能是N人群在O人群的排挤下北上的。总之,N人群在东南亚中南半岛上发展状况明显不如O人群,成为次要族群或寄人篱下附属于O人群。此后,O系黄种人逐渐占据中南半岛,并产生新的分化,这已经是2,5万年之前的事情了。
2万多年前在北部湾一带O1(M119)单倍群出现了,这是O(M175)这个超级男性大祖先的后代男性中,又形成的三个超级男性大祖先之一。另外两个是O3(M122)和O2(P31)。
我们必须清楚,每一个男性超级大祖先的出现,都意味着周围族群的悲剧,因为这是划时代的事情,周围族群将被逐渐淘汰,将被取而代之。这必然是一个血腥的过程。比如,O1、O2、O3这三个超级男性祖先本人和其直系后人,估计在上千年或几千年内,在扩大自己族群的同时,挤压着同期的各个O系远亲部落的生存空间,以至于几乎被灭绝。
在上百年,或几百年的时间段内,族群之间取而代之的事件很少见。但在跨度上千年或几千年的时间段,族群之间的取而代之事件就容易发生了。比如,在东南亚中南半岛上,这10万年内发生多少次族群之间的取而代之呢?先是6万年前D人群取代了丹人,然后是C人群取代了D人群,再然后是O人群取代了C人群。这是大跨度的族群取代,似乎屈指可数。实际上在D人群内部和C人群内部不知道又发生了多少次族群取代。比如,我们刚刚说过的O人群内部就存在这样的内部族群之间的取代(O人群取代NO人群,O1、O2、O3人群取代各自范围内的O人群)。幸运的是,O1、O2、O3这三个兄弟族群在相互火并的时候,出现了北上的机会,这样在中南半岛上的火并暂时告一个段落了。
可以说,大部分族群取代事件的发生,就是一次重要的基因进化或族群淘汰事件。这就是人类进化,虽然我们能侃侃而谈,但其细节我们还是情愿回避的。虽然A、B、C、D、O等单倍群的标志基因都是隔离有效基因的无影响的填充基因或其变异,但实际上,无影响的填充基因会变异,有效基因也会变异。正是这些有效基因的有益变异(有效基因的绝大部分变异都是有害变异)促使了人类的进化。如果是女性基因的有益变异,其会缓慢的影响族群,逐渐更多的有利于族群的发展。男性基因的有益变异可以更快的影响族群的发展,因为其优势基因的普及速度可以更快,一次或几次积累的男性有益基因变异,带来的基因优势如果可观,这意味着一个超级男性祖先诞生了,部落的原有平衡关系将被打破,周围部落被取而代之将成为历史命运。
第四纪晚更新世冰期时期,一些海拔不高的山也积雪冰封,使得积雪冰封的南岭等山脉成了黄种人向北进入长江流域的天然障碍,更主要的是南岭北部广大地区较低的冬季气温,不利于长期在热带生活的黄种人与已经长期生活在那里的土著竞争。
1万年前,冰期结束,东亚冰雪融化,气候变得温暖起来,长江流域土著的气候优势消失,黄种人竞争力相对大增,处于竞争的优势地位,逐渐取代当地土著成为历史趋势。
黄种人在中南半岛拥有着相对长江流域土著更先进的生产力,很快就开创了东亚全新的新石器时代,农业文明的星星之火在各地的黄种人中燃烧了起来,社会发展的火车头开动了。
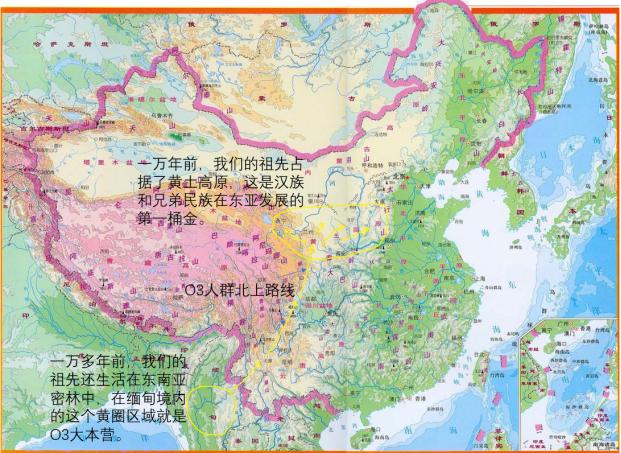
黄种人的北上分三条路线进行。内陆支黄种人O3沿着四川的几条大河(金沙江-雅砻江-大渡河)北上,来到了黄河上游地区,形成了汉藏(羌华)民族。
O3(M122)出现频率最高地区为中国云南省、印度东北部那加邦、缅甸的藏缅族群里,一些民族中出现频率为100%,如独龙族等,这是证明汉族起源于那里的基因学证据。这显示内陆支黄种人O3最初聚集在缅甸、中国云南省西部。
O3后来又分化为O3a(M324)、O3b[M164(O3a2)]、O3c[M159/Line1(O3a3)]、O3d[M7(O3a4)]、O3e[M134(O3a5)]。O3a分化出O3a1(M121)、O3d分化出O3d1(M113)、O3e分化出O3e1[M117/133(O3a5a)]。O3e1进一步分化出O3e1a(M162)。
每一次分化都意味着新的超级男性祖先的诞生,这意味着当时只有少数男性才留下了自己的直系后代。众多的其他男性只能是打酱油的身份,成为人类发展历史的匆匆过客。值得安慰的是,虽然没有直系后代,由于基因的重组遗传,其基因还是会有一点保留在自己族群的后人中的。
汉藏民族的一支形成了藏缅族群,他们创造了马家窑文化,以陇西为中心,西至河西走廊和青海省北部,北达宁夏南部,南抵四川省北部。马家窑文化是陕西仰韶文化的一种地方性的变体,年代为前3300~前2050年。齐家文化起源于中原之陕西龙山文化,随着不断向西发展,分布与马家窑文化地域重合,开始承袭马家窑文化的若干因素,地域性也就显得更加突出,实属陕西龙山文化的变体。齐家文化早期为公元前2000年,已普遍出现铜器,晚期年限可到商代。至于甘肃西部的火烧沟文化,则已进入比较发达的青铜时代,为前1900~前1700年。
藏缅族群不断南下,占领了原属棕色人种和兄弟民族百濮的很多地盘,并与之通婚,融合成现在的藏族人、羌族人、氐族人、彝族人、缅甸族人、蜀人、巴人(土家族)。
其中,藏族是秦汉时期藏缅族群中的两支唐旄发羌(仙女)南下进入青藏高原,与当地的棕色人种土著雅砻(猕猴)混血而形成的。现在的藏族人的遗传结构还能看到大量的D 型Y 染色体。而观察藏族的面貌特征,甚至在同一个村子中,也会看到两种截然不同的样式,有的人是挺拔的羌人的面容,有的人则是敦圆的棕色人种面貌,肤色的差异也很大。羌族含有部分D 型,但以O3 为主。另外,整个藏缅族群由于在黄河上游老家跟C融合的厉害或者因为后来匈奴、鲜卑、金山突厥、契丹、蒙古的影响,所以现在都含有10%以上的C血统。
O3在南亚语群体和苗瑶语群体中也存在。唯有O3e1是汉族才独有的(这很有可能是最后的或最近的一个超级祖先,毕竟进入成熟农耕文明社会以后,人口数量得以极大膨胀,基因结构渐趋稳定,任意某个男性再也难以传播占据明显优势的后代了,许多男性不再只是匆匆过客了,自己的直系后代也可以长期繁衍下去了),这个类型诞生于原始汉人的故乡黄土高原(甘南、宁夏、陕北),很可能是做了800年江山的周族的祖先帝喾-后稷(弃)。
汉藏民族另一支东进,形成原始汉人。公元前8000年,原始汉人逐渐散布整个黄土高原和华北平原西半部。原始汉人具有简单形式的颅顶缝、圆钝的眶形、突出的颧骨、低凹的鼻梁、浅的犬齿窝、低矮的鼻前棘以及比较扁平的面部;颅长中等兼高颅型、眶型中等偏低、中等的面宽和面高、阔鼻倾向、齿槽突颌以及中等的身高(平均约1.68米),接近现代黄种人的东亚(远东)类型,其中某些特征(如阔鼻、低眶及突颌等)更接近南亚类型,或可视之为继承了旧石器时代晚期智人的特征。

黄土高原景观图。黄土高原现在已经是千沟万壑了,森林覆盖率很低,但在汉族祖先来到黄土高原的时候,景观并非如此。那时的黄土高原大约50%的森林覆盖率,剩余的是草地。黄土高原西部的森林覆盖率会低于50%,以草地为主。汉族祖先之所以能在黄土高原获得大发展,这来源于黄土高原相对不那么潮湿的气候,在这里获得一定发展之后,才可以挺进相对潮湿一些的华北平原。在华北平原生活的汉族先民生产生活技术获得足够前进以后,才可以向较为潮湿的淮河以南发展。这就是汉族农耕文明地域扩张的基本模式。
中间支黄种人O2又称南亚(百濮)/澳斯特罗-亚细亚,形成于柬埔寨-泰国,并散布于整个中南半岛(除缅甸北部)、中国云南省东部、广西省和海南岛,今中国的佤-德昂-布朗、缅甸-泰国的孟族、柬埔寨的高棉族、越南的京族、马六甲族、尼科巴族、印度的蒙达族、斯里兰卡的维达人都是其后代。另外,百越民族中,泰国泰族的65%、中国壮族的35.95%也是O2的后代。O2后来又分化为O2a(M95)和O2b(SRY465),从O2a群体中又分化出O2a1(M88/111),O2b分化出O2b1(47z)。
中间支黄种人南亚(百濮)民族的一支南蛮(苗瑶),北上经过贵州、到达四川,接着沿着长江往下走,到了长江中游(以江汉平原为中心,南包洞庭湖平原、西尽三峡川东、北达豫南豫中、东抵鄱阳湖的广大地区),这就是历史上所谓的三苗,Y染色体是O2a*。后来汉藏部落O3d[M7(O3a4)]在华夏内战中落败混入南蛮成为其统治者,而南蛮的O2a1则脱离南蛮混入南部百越。南蛮先后创造了石门皂市下层文化(为前5000年)→大溪文化(前4400~前3300年)→屈家岭(湖北京山县)文化(前3000~前2000年)→湖北龙山文化(前2400年的龙泉三期,与中原龙山文化有较大区别)四种新石器文化。中间支的南亚(百濮)民族的其他支,没有随苗瑶支北上,而是继续留居中南半岛故乡。
沿海支黄种人O1称澳泰族群,线粒体单倍群B4a、B4b、M7b是其母系特征。O1后来又分成两大家族O1a(M101)和O1b(M50/103/110)。其分化和扩张分为三个阶段。
第一个阶段是旧石器时代的分化,首先是广西内陆的山地群体和北部湾沿海群体发生了差异。山地群体后来发展成为雒越(O1仅是统治阶层,土著可能为百濮),北部湾沿海群体沿中国海岸线迁徙而扩张开来,首先到达海南岛,形成黎族的祖先。1.4万多年前,北部湾沿海群体散布到广西东部(1万年前演化成仡佬族)、广东(侗傣族群祖先)和台湾(高山族祖先);稍后,北部湾沿海群体又有部分沿越南海岸线南下,后来发展成马来族群。
8000多年前,广东东部的某一个部族中开始萌发新石器文化(农业文明),并分三支迅速扩张开来。第一支向东北迁徙进人江西,形成干越;部分干越后来东迁进人浙江,形成了於越,在太湖平原和宁绍平原创造了灿烂的新石器文化。於越先后创造了浙江余姚河姆渡文化(前5000~前4000年)、太湖流域马家浜文化(分为马家浜和崧泽两期,为前4300~前3300年)、良渚文化(前3300~前2200年,与山东龙山文化有更密切的联系)、南京北阴阳营文化、安徽潜山薛家岗第三期文化(前3200~前3000年)。於越民族继而又从江苏进入山东,与当地的苗瑶、通古斯融合创造了先东夷文化。第二支渡海迁徙到台湾岛,形成了台湾平埔族的一部分。第三支在广东福建内部演化为闽越、南越。5000~3000年前,闽越向北分出了东瓯;南越向西分出了西瓯(西瓯和山地群体雒越融合形成了壮族)。广西东部的仡佬族祖先向西北迁徙到了贵州,建立了夜郎诸国,同化了当地的许多南亚语百濮族群,当今的仡央族群(包括仡佬、木佬、拉基、布央、普标、拉哈等语言, 分布在黔滇桂及越南北部)遗传结构可以看到明显的二元性。
澳泰族群分化扩张的第三个阶段是中华帝国时期的流散迁徙。闽越在汉武帝强制移民政策下,全部离开了福建,大部分到了江淮,逐渐同化于那里的汉族;小部分向广东迁移,并最终迁徙到贵州,演化成水族。广东的南越也大规模向西北迁徙,形成了今天的侗族。南越向西分出的西瓯和山地群体雒越融合形成壮族,唐朝时一部分继续西迁到西双版纳、南迁到老挝、泰国,征服并同化当地的孟高棉(百濮)土著,形成傣族/泰族。今壮族的17.9%、泰国泰族的10%、越南京族的7%、柬埔寨高棉族的8%、土家族的20%拥有M119。
澳泰族群北部湾沿海群体之一部,沿着越南海岸南下,来到马来半岛,征服当地的南亚百濮土著,形成南岛族群,接着向南面广阔的大洋进发。公元前3000年之后,扩散到了印尼、菲律宾。公元后又进一步扩散到非洲的马达加斯加、密克罗尼西亚[马绍尔、吉尔伯特(基里巴斯)、关岛查莫罗、加罗林群岛(波纳佩-雅普-特鲁克)、帕拉乌、瑙鲁]、美拉尼西亚[斐济、莫图、雅比姆、所罗门群岛、瓦努阿图]、波利尼西亚[新西兰毛利、乌维亚、萨摩亚、汤加、图瓦卢(艾利斯)、钮埃、库克群岛拉罗通加、社会群岛塔西提、土阿莫图、马克萨斯、夏威夷]。波利尼西亚南部群岛的棕种C含量浓厚。
黄种人沿海支和内陆支大规模来到北方以后,可能对当地的阿尔泰-棕种男人进行了杀戮和驱逐,但保留了阿尔泰-棕种女性以婚配。当历史进入到阶级社会,情况就发生了变化,奴隶的需求使得黄种人对遭遇到的棕种男人不再灭杀。那段时期黄种人才到达的边缘地区,比如青藏高原、蒙古高原、日本列岛,棕色人种的成分C和 D被大量地保留了下来。
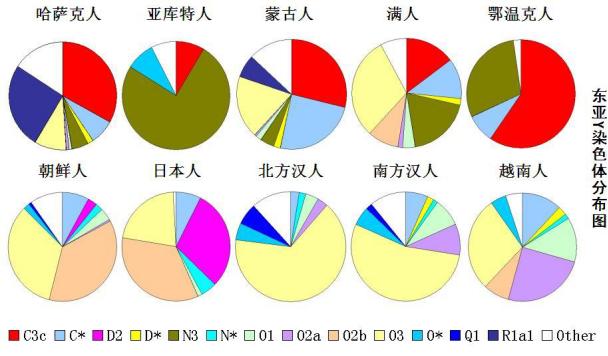
东亚部分族群Y染色体单倍群构成情况,东亚主要族群,比如朝鲜人、日本人、汉族人、越南人(虽然其地理上属于东南亚,但其人文等状况应归属于东亚)的主要Y染色体单倍群是O系,因此,可以说O系是黄种人的核心标志。汉族的核心标志是O3系,汉族是在O3系基础上融合其它族群,但主要是融合血缘关系最近的O系而形成。
,
-

- 张至顺道长口述一生修炼经历(104岁的张至顺7岁开始乞讨)
-
2023-08-07 02:33:50
-

- 潘粤明鬼吹灯系列应该先看哪个(潘粤明签了五部)
-
2023-08-07 02:31:34
-

- 王玉雯和王子奇结婚了吗(只是结婚的关系后)
-
2023-08-05 08:21:30
-

- 张艺兴爷爷什么人(原来我们似曾相识)
-
2023-08-05 08:19:15
-
- 张艺谋个人国籍(两个新身份曝光)
-
2023-08-05 08:17:01
-

- 张国立邓婕离婚(邓婕终于说出当年离婚的真相)
-
2023-08-05 08:14:46
-

- 侯明昊超级网剧寒武纪(寒武纪上演史上最萌杀手追爱记)
-
2023-08-05 08:12:32
-

- 张杰给谢娜庆祝40岁生日(张杰记错与谢娜结婚日)
-
2023-08-05 08:10:17
-

- 黑人说唱冷门rap(从一个郫县默默无闻的小青年)
-
2023-08-05 08:08:03
-

- 张靓颖 这就是爱情(张靓颖如果这就是爱情其实不是爱情)
-
2023-08-05 08:05:48
-

- 张耀扬现实老婆(56岁潇洒哥张耀扬)
-
2023-08-05 08:03:34
-

- 张翰王晓晨怎么回应(张翰演技遭质疑)
-
2023-08-05 08:01:19
-

- 王心凌最好听十首歌(王心凌最近很火的歌)
-
2023-08-04 10:11:10
-

- 张铁林谈谢园(谢园怎样一次恶搞让霸气威武的张铁林斯文扫地)
-
2023-08-04 10:08:56
-
- 王茜华模样大变(为拍戏打掉孩子)
-
2023-08-04 10:06:41
-

- 海清为中年演员发声(转头又给海清当女配)
-
2023-08-04 10:04:27
-

- 韩寒简介资料(韩寒的彪悍的游戏人生)
-
2023-08-04 10:02:12
-

- 何洁精修图原图对比(拍照人又矮又胖)
-
2023-08-04 09:59:58
-

- 佟磊说面(替罪羊佟磊因扮演了一个角色得罪冯小刚)
-
2023-08-04 09:57:43
-

- 张玉环为何被判27年(张玉环改判无罪)
-
2023-08-04 09:55:29
 蜜蜂仔真名叫什么 韦神妻子翁槿萱详细个人资料大揭秘
蜜蜂仔真名叫什么 韦神妻子翁槿萱详细个人资料大揭秘 靓妹仔失贞记洪晓芸个人资料
靓妹仔失贞记洪晓芸个人资料